Journal of Experimental Botany 63: 5337-5350 (2012)
Embryo growth, testa permeability, and endosperm weakening are major targets for the environmentally regulated inhibition of Lepidium sativum seed germination by myrigalone A [W][OA]
University of Freiburg, Faculty of Biology, Institute for Biology II, Botany / Plant Physiology, Schänzlestr. 1, D-79104 Freiburg, German (AV, KG, KO, DJ, GLM)
School of Biological Sciences, Plant Molecular Science, Royal Holloway, University of London, Egham, Surrey TW20 0EX, Uk (AV, KG, GLM);
Web: 'The Seed Biology Place' - www.seedbiology.eu
Department of Plant Physiology, Warsaw University of Life Sciences-SGGW, 15 Nowoursynowska 159, 02-776, Warsaw, Poland (KO)
Laboratory of Growth Regulators, Faculty of Science, Palacky University and Institute of Experimental Botany AS CR, v.v.i., Šlechtitelu 11, CZ-783 71, Olomouc, Czech Republic (DT, VT, TU, MS)
Centre of the Region Haná for Biotechnological and Agricultural Research, Faculty of Science, Palacky University, Šlechtitelu 11, CZ-783 71, Olomouc, Czech Republic (MS)
Received March 10 2012; Revised June 11 2012; Accepted June 15 2012
Advance Acess publication July 21 2012
DOI 10.1093/jxb/ers197
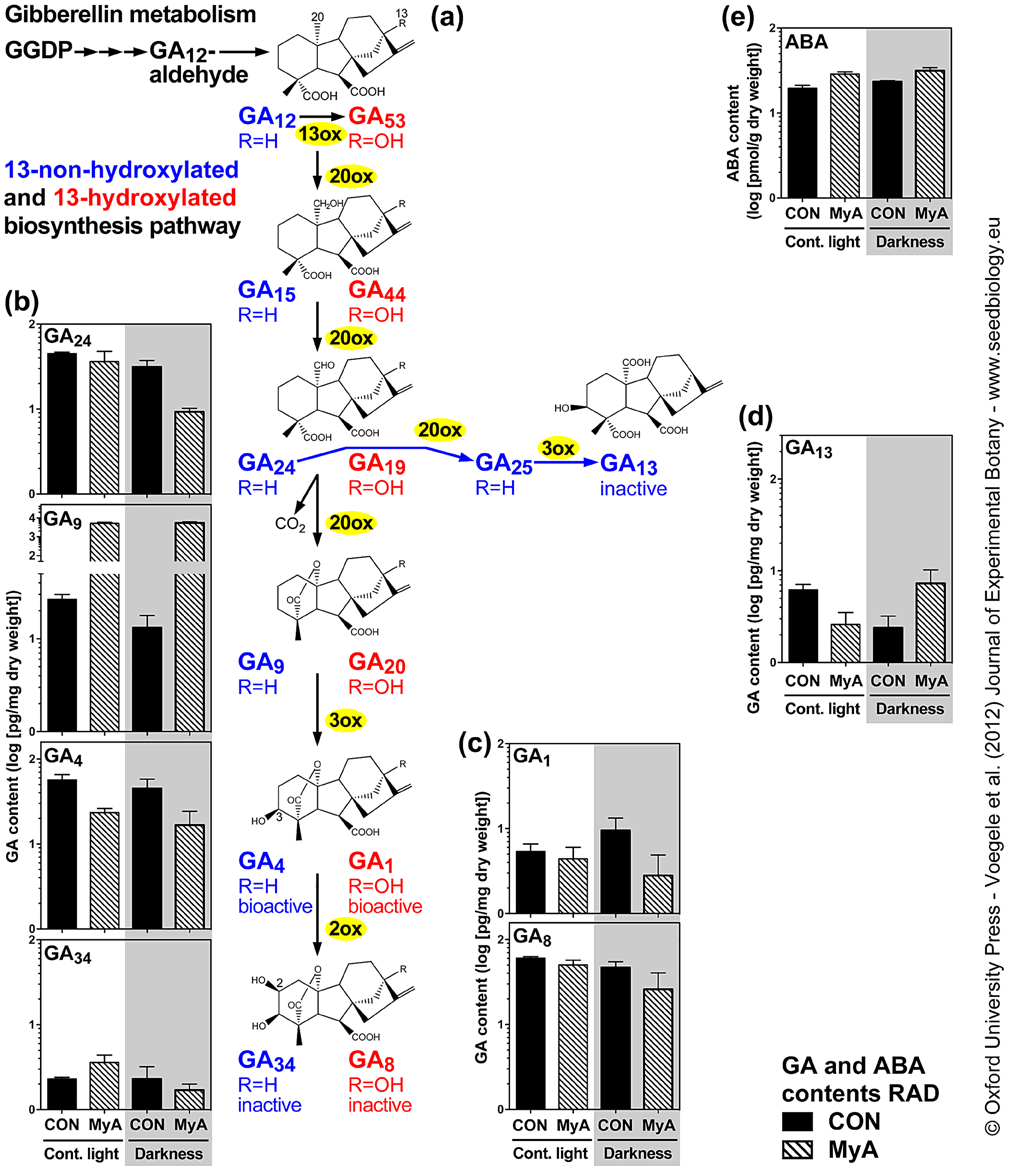
Figure S4. The tissue-specific effect of MyA on gibberellin (GA) and ABA metabolism during the germination ofLepidium sativum seeds incubated in continuous light or darkness without (CON) or with 5x10-4 M MyA added.
(a) The 13-non-hydroxylated (blue) and the 13-hydroxylated (red) GA biosynthesis and inactivation pathways and important metabolites detected in L. sativum seeds.
(b-d) Contents of bioactive gibberellins such as GA1 and GA4, and inactive forms such as GA24, GA9, GA34, GA8, GA13 and of ABA (e) quantified in RAD extracted from CON and MyA-treated seeds incubated for 15h. For comparison to darkness (this work), values for continuous light (Oracz et al. 2012) are presented.
Note that metabolite contents are expressed in log scale as mean values ± SE of four biological replicates.
| Article in PDF format (1.5 MB) Supplementary data file (2 MB) |
|
|
|
The Seed Biology Place |
Webdesign Gerhard Leubner 2000 |