Tansley Review - New Phytologist 171: 501-523 (2006)
Seed dormancy and the control of germination
(2) Institut für Biologie II (Botanik/Pflanzenphysiologie), Albert-Ludwigs-Universität Freiburg, Schänzlestr. 1, D-79104 Freiburg i. Br., Germany, Web: 'The Seed Biology Place' http://www.seedbiology.de
Received February 23, 2006; accepted March 27, 2006; published
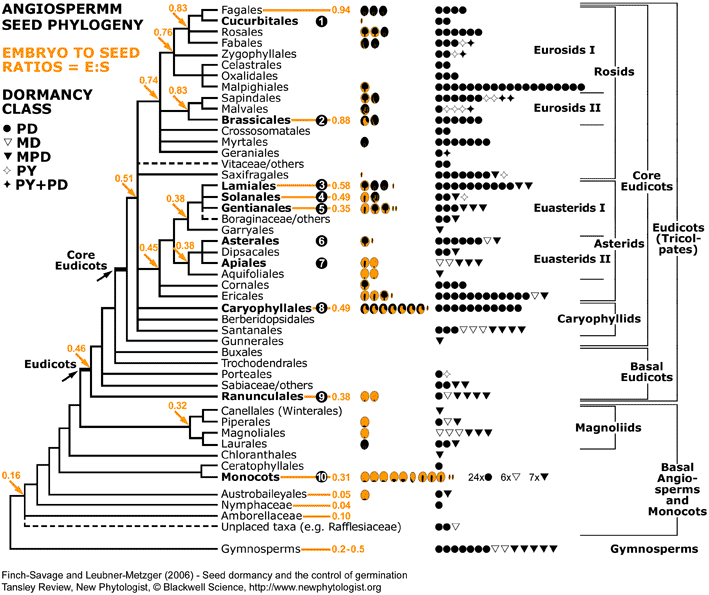
Figure 4. Angiosperm seed evolution depicted in a phylogenetic tree of the Angiosperm Phylogeny Group II (2003). For each family, pictographs of the seed type and symbols for the dormancy class are placed next to the corresponding clade (see Fig. 3 for symbols and data source). The number of pictographs or symbols is equal to the number of families with the corresponding seed type or dormancy class, respectively. Numbers in the phylogenetic tree represent "embryo to seed" (E:S) ratios expressed as GSL (generalized least squares) values from Forbis et al. (2002, personal communication). Clades with experimental evidence for endosperm-limited germination and endosperm weakening are in bold and numbered (1) to (10). The experimental evidence can be summarised as follows: Rosid clade: (1) Cucurbitaceae: Cucumis C
| Article in PDF format (2 MB) |
|
|
|
The Seed Biology Place
|
Webdesign Gerhard Leubner 2000
|