Tansley Review - New Phytologist 171: 501-523 (2006)
Seed dormancy and the control of germination
(2) Institut für Biologie II (Botanik/Pflanzenphysiologie), Albert-Ludwigs-Universität Freiburg, Schänzlestr. 1, D-79104 Freiburg i. Br., Germany, Web: 'The Seed Biology Place' http://www.seedbiology.de
Received February 23, 2006; accepted March 27, 2006
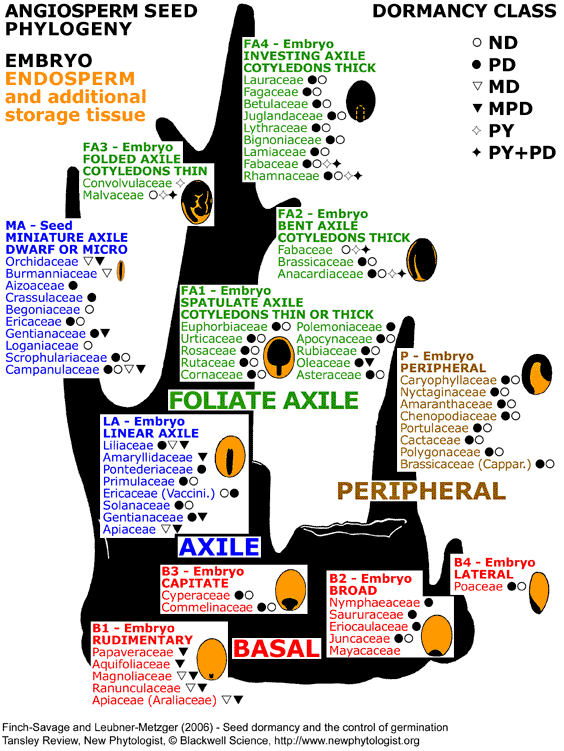
Figure 3. Seed phylogenetic tree based on the internal morphology of embryo and endosperm in mature angiosperm seeds. Martin (1946) investigated the embryo (form, size, position) and endosperm (plus additional storage tissue) in 1287 species and proposed seed types (B1 to B4, LA, P, MA, FA1 to FA4). Seed types with abundant endosperm (orange) and a tiny embryo (black) are basal (B1, B2, B3, B4). In the more advanced endospermic LA-type seeds, the embryo is linear axile. From this developed the FA-type seeds (FA1, FA2, FA3, FA4) where the embryo is foliate axile and, depending on the subtype, differs in shape and occupies more or less the entire seed. Mature FA-type seeds have little or no endosperm, and the nutrients are stored in the cotyledons. The evolution of whole seed size (e.g. dwarf seeds, MA) is beyond the scope of this review and is discussed elsewhere (e.g. Baskin & Baskin, 1998; Baskin & Baskin, 2005; Fenner & Thompson, 2005). Seed dormancy classes are indicated next to each family name: non-dormancy (ND
| Article in PDF format (2 MB) |
|
|
|
The Seed Biology Place
|
Webdesign Gerhard Leubner 2000
|