Tansley Review - New Phytologist 171: 501-523 (2006)
Seed dormancy and the control of germination
(2) Institut für Biologie II (Botanik/Pflanzenphysiologie), Albert-Ludwigs-Universität Freiburg, Schänzlestr. 1, D-79104 Freiburg i. Br., Germany, Web: 'The Seed Biology Place' http://www.seedbiology.de
Received February 23, 2006; accepted March 27, 2006
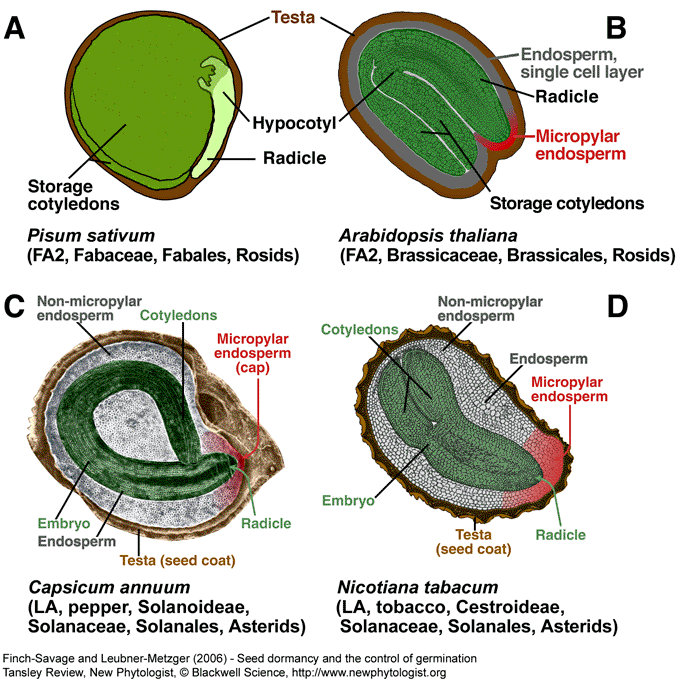
Figure 1. Biodiversity of the structure of mature seeds of angiosperms and the importance of the seed covering layers. The diploid embryo is surrounded by two covering layers: the triploid (most species) endosperm (nutritive tissue, mostly living cells) and the diploid testa (seed coat, maternal tissue, mostly dead cells). In several species the endosperm is completely obliterated during seed development and the nutrients are translocated to storage cotyledons. Mature seeds of pea (A, without endosperm) and A. thaliana (B, single cell layer of endosperm) are characterized by embryos with storage cotyledons. The micropylar endosperm (several cell layers) is known to be a germination constraint of Solanaceae seeds (C,D). FA2 and LA are seed types (see Fig. 3). (C) Modified based on Watkins and Cantliffe (1983), reprint with permission, © American Society of Plant Biologists. (A,B,D) “The Seed Biology Place” - http://www.seedbiology.de
| Article in PDF format (2 MB) |
|
|
|
The Seed Biology Place
|
Webdesign Gerhard Leubner 2000
|